A Hearty Introduction:
Before we get going, here are some exciting heart facts to start your day with: In case your momma never told you, hearts have 2 types of chambers: atria and ventricles. Atria are where blood enters the heart and ventricles pump the blood out of the heart. Hearts take in oxygen poor blood, pump it through the pulmonary circuit (lungs/gills) where it gets oxygenated, and then they pump it out to the rest of the body. There are many many small veins (take blood to the heart) and arteries (carry it away) which connect the heart to the pulmonary circuit and keep it all running right. Blood pressure has to be carefully balanced in all tubes so that flow pressure is maintained and blood keeps moving, but not so fast as to explode certain areas or capillaries. One might say this careful and complex hydrostatic wiring might be easily selectable in an evolutionary scenario, but there's no getting around the fact that there's a very complicated and highly balanced network of biological fluid mechanics going on inside a heart than most artificial heart engineers probably care to think about.
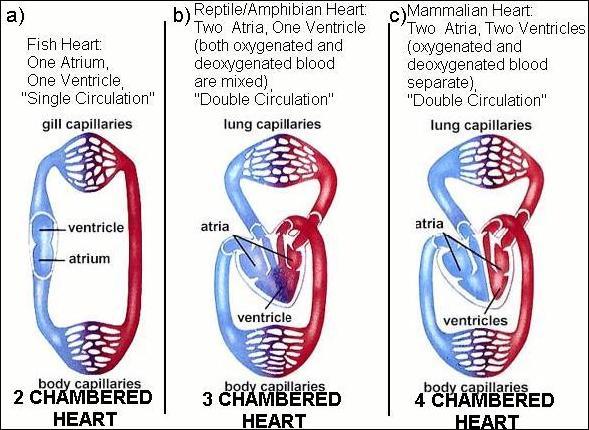
Given all this, there are 3 basic ways to make a heart found in animals: a 2 chambered heart, a 3 chambered heart, and a 4 chambered heart. Fish have 2 chambers, one atrium and one ventricle. Amphibians and reptiles have 3 chambers: 2 atria and a ventricle. Crocodiles are the one reptilian exception, as they have 4 chambers (2 atria, 2 ventricles). Birds and mammals have 4 chambers (2 atria and 2 ventricles).
Differences between the hearts:
The fish heart (figure 1a) is much different than the amphibian/reptile/bird/mammal heart (figures 1b and c). Hearts are very complex--they're not just a bunch of random arteries and veins connecting tissue. Fish hearts simply draw in deoxygenated blood in a single atrium, and pump it out through a ventricle. This system is termed "single circulation", as blood enters the heart, gets pumped through the gills and out to the body, Blood pressure is low for oxygenated blood leaving the gills.
3 and 4 chambered hearts have a pulmonary circuit (pathways taking blood from heart to lung and back to heart) that is very complex and must be set up such that blood can travel from the heart to become oxygenated in the lungs and then be properly pumped back the heart and out to the body. The 3 (and 4) chambered heart has "double circulation" (figure 1b and c) and is quite different from "single circulation" (figure 1a) of fishes.
"Double circulation" has an interior circuit within the heart--blood enters the heart, leaves the heart and gets oxygenated, enters the heart again, and then gets pumped out to the body. Because "Double circulation" allows oxygenated blood to be pumped back into the heart before going out to the body, it pumps blood with much more pressure and much more vigorously than "single circulation".
Figure 1:
The Above diagram adapted from "
http://gened.emc.maricopa.edu/bio/bio181/BIOBK/BioBookcircSYS.html#Vertebrate Vascular Systems which was said to be adapted from
http://www.biosci.uga.edu/almanac/bio_104/notes/may_7.html and bears a striking resemblance to the diagrams from Campbell's "Biology" 4th edition pg. 822.
Though the 4 chambered heart has 2 atrium-ventricle pairs, both pairs do not do the same thing. There are 4 steps involved with blood entering the heart: 1) oxygen poor blood enters the first atrium. 2) oxygen poor blood is fed to the first ventricle, which pumps it out to the pulmonary circuit (lungs) where it is enriched in oxygen. 3) Oxygen rich blood just leaving the lungs is pumped back into the second atria. 4) Oxygen rich blood is then fed to the second ventricle, which pumps the oxygen rich blood out of the heart and back into the body for usage.
The 4 chambered heart differs from the 3 chambered heart in that it keeps oxygenated blood completely separate from de-oxygnated blood, because there is one ventricle for deoxgynated blood and one for oxygenated blood. In the 3 chambered heart, a single ventricle pumps both out of the heart, and there is some mixing between fresh and old blood. The 2 ventricle-4 chamber heart prevents mixing allows the blood leaving the heart to have far more oxygen than it would otherwise. This is good for enhancing the more fast paced lifestyle that birds and mammals tend to have, giving an advantage to having a 4 chambered heart.
Problems with evolving the hearts:
Getting a heart, period: Evolving a 2 chambered heart in the first place is very difficult because the circulatory system is irreducibly complex. At least 3 subsystems are necessary: 1) an organ for enriching the hemophlegm (blood) with oxygen (lungs/gills), 2) a complex network of closed tubes to carry the energy-rich blood to the body (veins and arteries), and 3) a pumping mechanism (heart) to transport energy-rich fluid throughout the body. One common way an evolutionist might try to get around problems associated with irreducible complexity might be to imagine scenarios where some or all of the subsystems could originate in a freestanding manner, functional on their own. However in this case what good is a closed tube network without a pumping mechanism to transport fluid, and what good is a pump without the fluid or the tubing? Oxygen exchange occurs in many organisms through the skin without a chordate-like circulatory system, but what is the advantage for such an organism to randomly mutate itself a single oxygen exchange organ (the lungs/gills)? Even so, once a single oxygen exchange organ was in place, it would need the very transport network provided by the heart, veins and arteries. Scenarios attempting the circulatory system in a step by step manner would fail and not take into account the overall complexity of the system.
Even if the subsystems of the circulatory system could be evolved on their own, evolving a freestanding 2 chambered heart on its own would be very difficult, even if it didn't need other components of the circulatory system to be useful. A human heart is an 11 ounce pumping machine the size of a human fist which beats over 2 billion times and pumps over 100 million gallons of blood over the course of the average human lifetime. It is primarily a shell with a carefully balanced interior network of holes and valves which keep fluid constantly flowing in, out, and in the right direction everywhere in between. A large number of fluid-directing parts as well as a very strong and properly and complexly shaped pumping muscle are necessary for it to function. The positioning of the parts of a heart are a good example of
specified complexity. For this reason, the heart, as well as the entire circulatory system, are unevolvable in a step by step manner.
Getting a 3 chambered heart from a 2 chambered heart: Given a 2 chambered heart, experts do not know when, how, or in what lineage the alleged transition from the 2 chamber fish heart to the 3 chambered amphibian heart took place, mainly because this is a very difficult transition to even imagine. A 3 chambered heart has "double circulation" and is irreducibly complex with respect to "double circulation". 2 chambered hearts of fish have "single circulation" and the basic design is very different from a 3 chambered heart.
A vastly oversimplified explanation of the evolution of the heart might be to say that by simply duplicating the some or all of the chambers of a 2 chambered heart, one could easily evolve a functional 3 or 4 chambered heart. Of course the way the evolutionary story goes, fish turned into amphibians which turned into reptiles which became mammals and birds, so the heart is said to have evolved from 2 --> 3 --> 4 chambers. A direct 2 --> 4 chamber transition is never thought to have taken place and would have even more difficulties than a 2 --> 3 chamber transition, so we will focus on the 2 --> 3 chamber transition. However, in reality the changes that would need to take place for this transition are far more than merely the duplication of one or more chambers. It could not happen in a step-by-step manner where intermediates are functional.
The 2 chamber heart --> 3 chamber heart transition requires much more than the duplication of an atrium, because the interior circuit causing the "double circulation" of the 3 chambered heart must also be created. Duplicating the atrium without a closed circulatory network for "double circulation" would cause the heart to suck nothing but interstitial fluid out of the body. "Double circulation" only works when there is a loop feeding from a ventricle back to the heart, and back to a ventricle. Thus, both the loop and the new atrium are necessary for a 3 chambered heart to function. And though the loop (interior circuit) sounds simple, it really must be a complex tube network with valves in the right places to keep fluid flowing properly.
Single circulation hearts pump blood directly through the gas exchange organ and out to the body (figure 1a). Double circulation pumps blood to the lung/gills through an "interior circuit" loop and then back to the heart before going out to the body (figure 1b and c). And if this new loop doesn't connect with the gas exchange organ, then the new loop is functionless and useless. The easiest way to make this transition happen is probably to have the vein leaving the gas exchange organ feedback to become an artery feeding back into the heart. Of course this means the new vein-artery simultaneously needs the proper valves so it can function like an actual part of the heart itself. Probably of most importance is the fact that there is now no vein leaving the heart and pumping blood back out to the body. In other words, to create this interior circuit of double circulation, a new vein must be created and blood flow out of the heart completely rewired to the rest of the body.
Even if gas exchange organ could be bypassed (making it a less complex, though functionless and not selectively advantageous circuit), the main problem with going from single circulation to double circulation is that somehow this new circuit has to wire itself to the lungs. Either way, the vein leaving the heart must somehow also become an artery fed back into the heart through a new functional atrium and then a new vein must be created so that blood leaving the heart still gets out to the rest of the body through the circulatory network.
Finally, the heart muscle has to adapt to all of these changes, especially such that beating can occur to pump through the new atrium and associated fluid pressure changes.
In other words 4 primary changes are needed to go from single to double circulation:
The duplication of the atrium such that fluid transport through new atrium is functional
A conversion of the vein leaving the heart into an artery at the other end such that it is fed back into the heart.
A complete rewiring of how blood finally leaves the heart and goes out to the rest of the body (the creation of a new vein and rewiring).
Modification of the heart muscle to beat properly and accommodate the additional atrium and fluid pressure changes associated with the rewiring.
If any of these steps are missing, double circulation won't work. And this says nothing about the many valves and other smaller veins and arteries associated with double circulation which characterize true hearts as well changes needed in the pumping mechanism of the heart muscle to accommodate a completely new atrium and fluid-pressure balance.
The transition from 2 to 3 chambers requires a change from single to double circulation which involves at least 4 major simultaneous changes including the complete rewiring of how blood leaves the heart to the rest of the body. Many more minor simultaneous changes associated with mechanics of proper fluid transport would also be necessary. It not possible for double circulation system to evolve from a single circulation heart system in a Darwinian step-by-step manner because too many changes are necessary, making the 3 chambered heart unevolvable from a 2 chambered heart.
Getting a 4 chambered heart: With respect to hearts with "double circulation", the 4 chamber mammalian heart probably isn't irreducibly complex. Going from 3 to 4 chambers really doesn't look all that complicated (of course they're still very different and this is totally oversimplified, but I'm only talking about basic organ design). Basically, the single ventricle in the 3-chambered heart is split into 2 chambers in the 4 chambered heart, making 2 ventricles instead of one. A "double circulation" heart could work with 3 chambers--and it does in reptiles and amphibians.
The human heart has 2 atrium-ventricle pairs, which beat in succession something like pistons in a car. Only one ventricle is really needed to pump the blood. But just like an 8 cylinder engine put out a lot more horsepower than a 4 cylinder, so does a 4 chamber (2 pairs) heart have a much more power to supply the body with energy and oxygen-rich blood than a 3 chamber (1 ventricle) heart. There is a huge advantage to having a 4 chamber heart. Taking away the additional capillary complexities associated with suddenly having a second ventricle, one might be able to argue that the 3 to 4 chambered heart transition isn't difficult to imagine, relatively speaking, as all one has to do is note the strong advantage of separating oxygenated and de-oxygenated blood and then divide the ventricle in half. Of course the key word here is "imagine".
Irreducible complexity is a real phenomena, and it can be analyzed, and so in some cases it might not exist, and in the case of the 4-chambered heart case it probably doesn't. Though the 4 chambered heart may not be irreducibly complex with respect to "double circulation", it might still be the result of intelligent design and not evolution, and irreducible complexity doesn't have to exist in all instances for it to exist in some. Overall, regardless of chambers, the heart has had much design put into it. And the complex double-circulatory heart bears strong marks of intelligent design.
Getting a bird heart: Finally, the evolutionary origin of the bird heart-lung system is puzzling for the dino-bird hypothesis because "no lung in any other vertebrate species is known which in any way approaches the avian system." (Denton (1998), pg 361). Molecular biologist Michael Denton had the following to say about the evolutionary origin of the bird-heart-lung system:Just how such a different respiratory system could have evolved gradually from the standard vertebrate design without some sort of direction is, again, very difficult to envisage, especially bearing in mind that the maintenance of respiratory function is absolutely vital to the life of the organism. Moreover, the unique function and form of the avian lung necessitates a number of additional unique adaptations during avian development. As H. R. Dunker, one of the world’s authorities in this field, explains, because first, the avian lung is fixed rigidly to the body wall and cannot therefore expand in volume and, second, because the small diameter of the lung capillaries and the resulting high surface tension of any liquid within them, the avian lung cannot be inflated out of a collapsed state as happens in all other vertebrates after birth. In birds, aeration of the lung must occur gradually and starts three to four days before hatching with a filling of the main bronchi, air sacs, and parabronchi with air. Only after the main air ducts are already filled with air does the final development of the lung, and particularly the growth of the air capillary network, take place. The air capillaries are never collapsed as are the alveoli of other vertebrate species; rather, as they grow into the lung tissue, the parabronchi are from the beginning open tubes filled with either air or fluid (Denton (1998), 361)Denton goes on to say that, "The avian lung brings us very close to answering Darwin’s challenge: ‘If it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down.’
One respiratory physiology expert, John Ruben, critiquing this evolutionary scenario noted that, "a transition from a crocodilian to a bird lung would be impossible, because the transitional animal would have a life-threatening hernia or hole in its diaphragm ... It seems clear that a bird’s radically different system of breathing, in which air is continuously drawn through its lungs, could not have evolved from the hepatic-piston system we see in this theropod dinosaur." (Gibbons, 1997 B, pg. 1230)
Plus, we're also dealing with convergent evolution here. The 4 chamber system had to have evolved twice (and also in crocodiles), if it evolved at all, because mammals and birds are allegedly from very different reptilian stock. It isn't just a neat case of common ancestry being clear cut by characteristics.
There are many other animals with hearts--some worms have like 5 or 6 hearts. These heats all probably evolved independently however. So, there is not a nice neat Darwinian tree with respect to hearts. Common design better explains this, because odds are low you would come up with a similar structure through chance evolution alone.
Summary and conclusion:
Fish: 2 chambers, Amphibians and reptiles: 3 chambers, Mammals and birds: 4 chambers. 3 and 4 chambered hearts are similar because they both have an interior circuit or "double circulation". This is very different from the 2 chambered heart which has "single circulation". Going from a 2 to a 3 chambered heart requires A LOT more than the mere duplication of a chamber, but a COMPLETE reworking of the veins and arteries surrounding the heart, interior valves of the heart, and the creation of this interior circuit which can properly interact with the lungs. Plus I have it on good authority that evolutionists don't know how the 2 to 3 chamber transition took place. Going from 3 to 4 might be more possible in a remote theoretical sense, though there are surely many less important complications. Going from 2 to 4 requires a vast more change than mere duplication of atria and ventricles. It might be remotely possible to go from a 3 chambered heart to a 4 chambered heart but it is not likely that a 2 chambered heart could evolve into a 3 or 4 chambered heart.
























